
Professor Emeritus
Ph.D. Plant Physiology University College of Wales
B.S. Botany University College of Wales
Organisms respond to environmental signals including light, temperature, and water supply and endogenous signals such as hormones, metabolites and other regulatory molecules. Cells transduce these signals into a specific response using signal transduction pathways. Our research focuses on the mechanisms of hormonal signaling in plant cells. Using the cereal aleurone as a model system, we research the signal transduction pathway regulated by the antagonistic plant hormones gibberellic acid (GA) and abscisic acid (ABA).
Signal transduction in plants
Organisms respond to a multiplicity of environmental signals including light, temperature, and water supply and endogenous signals such as hormones, metabolites and other regulatory molecules. Cells transduce these signals into a specific response using signal transduction pathways. Our research focuses on the mechanisms of hormonal signaling in plant cells. Using the cereal aleurone as a model system, we are unraveling the signal transduction pathway regulated by the antagonistic plant hormones gibberellic acid (GA) and abscisic acid (ABA).
In response to GA, the aleurone cell synthesizes and secretes a variety of hydrolytic enzymes that are essential for germination. In addition, aleurone cells exposed to GA undergo numerous physiological responses including a drop in cytoplasmic and vacuolar pH, an increase in cytosolic Ca2+ concentration and increases in the abundance of several other molecules known to play a role in signal transduction. GA-treatment is accompanied by morphological changes that cause the numerous protein-filled vacuoles of the aleurone cell to fuse to form one large central vacuole. Treatment of aleurone cells with ABA inhibits all of these GA-induced responses.
Recent efforts in the laboratory have focused on the events that lead to programmed cell death in the aleurone. Treatment with GA causes aleurone cells to undergo a series of uncharacterized events that lead to cell death. Whereas aleurone cells die within 6 days of exposure to GA, cells treated with ABA do not die for several months. Aleurone cell death is correlated with a loss in nuclear DNA and the activation of proteases and nucleases. The process of cell death can be accelerated by exposing GA-treated aleurone cells to reactive oxygen species or by treatments that increase the intercellular production of hydrogen peroxide. ABA-treated aleurone cells are much less susceptible to attack by reactive oxygen species than GA-treated cells. We are currently using a variety of techniques to characterize the morphological, biochemical and molecular aspects of cell death in the aleurone cell. Our long term goal is to unravel the mechanisms of hormone action in plant cells using the barley aleurone cell as a model system.
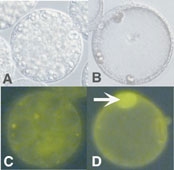
ABA-treated Aleurone cellshave many vacuoles (A), don’t stain with TUNEL (C).
GA-treated cells have one large vacuole (B) and stain with TUNEL (arrow in D), indicating degraded DNA.
Ultrastructure of barley (cv. Himalaya) aleurone layer cells illustrating their high storage oil content.
(a) Light microscopy of chemically fixed and plastic-embedded aleurone cells stained with periodic acid–Schiff reagent/aniline blue-black (PAS/ABB) showing large protein storage vacuoles (PSV, arrowhead) and prominent nucleus (N). Other organelles are poorly resolved at this magnification (approximately ×500).
(b) A polyethylene glycol-embedded thick section of aleurone layer and starchy endosperm (ST) from grains that have been centrifuged (Jones, 1969b). Lipids are stained black with Sudan IV and localize to the lower (centrifugal) ends of all aleurone cells. No lipid staining is visible in the starchy endosperm (approximately ×100).
(c) Chemically fixed and plastic-embedded centrifuged aleurone cells. Sections were stained with PAS/ABB. Organelles below the black line are oil bodies; PSV are located centripetally in the cell (approximately ×400).
(d) EM image of a section through a chemically fixed centrifuged cell. Small (approximately 0.1-µm-diameter) oil bodies (OB) are visible in the lower (centrifugal) half of the cell (approximately ×1500).
Fath, A, Bethke, P.C., Jones, RL 2001. Enzymes that metabolize reactive oxygen species are down regulated prior to GA-induced programmed cell death in barley aleurone. Plant Physiol, 126, 156-166.
Bethke, P.C., Fath, A. Jones, R.L. 2001 Regulation of viability and cell death by hormones in cereal aleurone. Int J Plant Phys 158, 429-438.
Lonsdale, J.E., McDonald, K.L., Jones, R.L. 2001 Microwave polymerization in thin layers of LR white allows selection of specimens for immunogold labeling. In: Microwave Techniques and Protocols, R.T. Giberson and R. S. Demaree Jr, eds., Humana Press (Totowa, NJ). p 139.
Fath, A., Bethke, P.C., Belligni, M.V., Spiegel Y.N., Jones, R.L. 2001 Signaling in the Cereal aleueone: Hormones, reactive oxygen and cell death. New Phytol 151, 99-109
Fath, A., Bethke, P.B., Belligni, M., Jones, R.L. 2002 Active oxygen and cell death in cereal aleurone. J. Exp Bot 53, 1273-1282.
Bethke, P.C., Fath, A., Spiegel, Y.N., Hwang, Y., Jones, R.L. 2002. Abscisic acid, gibberellin and cell viability in cereal aleurone. Euphytica, 126, 3-11.
Belligni,M., Fath,A., Bethke, P., Lamattina, L., Jones, R. 2002 Nitric oxide acts as an antioxidant and delays programmed cell death in barley aleurone layers. Plant Physiol 129, 1642-1650.
Zhang L, Liu BF, Liang S, Jones RL, Lu YT. 2002 Molecular and biochemical characterization of a calcium/calmodulin-binding protein kinase from rice. Biochem J. 2002 Nov 15;368(Pt 1):145-57.
Hwang YS, Bethke PC, Gubler F, Jones RL. 2003 cPrG-HCl a potential H+/Cl- symporter prevents acidification of storage vacuoles in aleurone cells and inhibits GA-dependent hydrolysis of storage protein and phytate. Plant J. 2003 Jul;35(2):154-63.
Bethke PC, Badger MR, Jones RL. 2004 Apoplastic synthesis of nitric oxide by plant tissues. Plant Cell. 2004 Feb;16(2):332-41. Epub 2004 Jan 23.
Bethke PC, Gubler F, Jacobsen JV, Jones RL. Dormancy of Arabidopsis seeds and barley grains can be broken by nitric oxide. Planta. 2004 Sep;219(5):847-55. Epub 2004 May 6.
Hwang YS, Bethke PC, Cheong YH, Chang HS, Zhu T, Jones RL. 2005 A gibberellin-regulated calcineurin B in rice localizes to the tonoplast and is implicated in vacuole function. Plant Physiol. 2005 Jul;138(3):1347-58. Epub 2005 Jun 24
Bethke PC, Libourel IG, Reinohl V, Jones RL. Sodium nitroprusside, cyanide, nitrite, and nitrate break Arabidopsis seed dormancy in a nitric oxide-dependent manner. Planta. 2005 Sep 3;:1-8
Libourel IG, Bethke PC, De Michele R, Jones RL. Nitric oxide gas stimulates germination of dormant Arabidopsis seeds: use of a flow-through apparatus for delivery of nitric oxide. Planta. 2005 Sep 17;:1-8
Eastmond PJ, Jones RL. 2005 Hormonal regulation of gluconeogenesis in cereal aleurone is strongly cultivar-dependent and gibberellin action involves SLENDER1 but not GAMYB. Plant J. 2005 Nov;44(3):483-93.
Sarath G, Bethke PC, Jones R, Baird LM, Hou G, Mitchell RB. 2005 Nitric oxide accelerates seed germination in warm-season grasses. Planta. 2005 Dec 21;:1-11.
Bethke PC, Libourel IG, Jones RL. 2006 Nitric oxide reduces seed dormancy in Arabidopsis. J Exp Bot. 2006;57(3):517-26. Epub 2005 Dec 23.
Bethke PC, Hwang YS, Zhu T, Jones RL. 2006 Global Patterns of Gene Expression in the Aleurone of Wild-Type and dwarf1 Mutant Rice. Plant Physiol. 2006 Feb;140(2):484-98. Epub 2005 Dec 29 Rec
24 - Freshman Seminar
99 - Supervised Independent Study and Research
290 - Graduate Seminar

Russell Jones
Berkeley, California 94720